

Cours abrégé de biologie végétale
Bonjour à tous, suite au topic de « » sur la biologie végétale je me suis rendu compte qu’il n’y avait pas vraiment de guide spécialement dédié à cette question.
Préambule :
Il existe plusieurs types de cellules dans un végétal :
- les cellules chlorophylliennes
- les cellules des poils absorbants
- les cellules des vaisseaux : Xylème (cellules mortes dont il ne reste que la paroi lignifiée) qui conduit la sève brute contenant les nutriments bruts et la plus grande partie de l’eau depuis les racines jusque dans les tiges et les feuilles, Phloème (Cellules semi-vivantes qui ont besoin de « cellules jumelles » qui leur permettent de rester en demi-vie) qui conduisent la sève élaborée contenant les sucres, les hormones, et d’autres molécules depuis les feuilles jusqu’à toutes les parties du végétal)
- les autres cellules (les tissus de soutient, celles du cœur des tiges et des racines, les réserves, etc…)
Elles possèdent toutes une paroi et certaines, les chlorophylliennes possèdent des chloroplastes qui sont le siège de la photosynthèse:
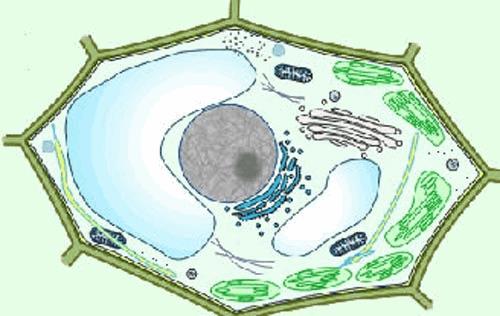
Nous verrons en plans assez larges les grands aspects de la biologie des plantes thérophytes (plantes qui ne survivent à la saison hivernale qu’à l’état de graine).
I. Le port du végétal :
Le port correspond à la façon dont pousse lé végétal, la forme qu’il va prendre en grandissant.
Voici les formes les plus classiques :
Port en boule ou buissonnant

Port arborescent (comme les sapins et notre plante)

Le port est déterminé par la concentration en Auxine (hormone végétale, voir plus bas) au niveau des bourgeons, cette différence de concentration détermine quel bourgeon sera le (ou les dans le cas du port en boule) dominant (qui grandira le plus vite).
Ensuite, le système racinaire de notre plante est de type pivot :
Et c’est pour ça que les pots les mieux adaptés sont des pots assez haut mais pas nécessairement très larges, le mieux étant des pots comme ceux utilisés pour les rosiers car ils ont un système racinaire du même type.
II. Les apex :
L’apex est la partie de la plante qui comprend la plupart des méristèmes, là où les cellules se divisent et où elles se différencient pour finalement donner les futures tiges et feuilles.
En général, on appelle apex le bourgeon principal qui se trouve au sommet de la plante mais en fait il y a un apex à la base de chaque feuille mais leur développement est inhibé par l’apex principal.
II.1. Les apex caulinaires :
L’apex caulinaire apical est celui qui se trouve au somment de la tige, le plus haut dans la plante lorsqu’elle n’est pas taillée.
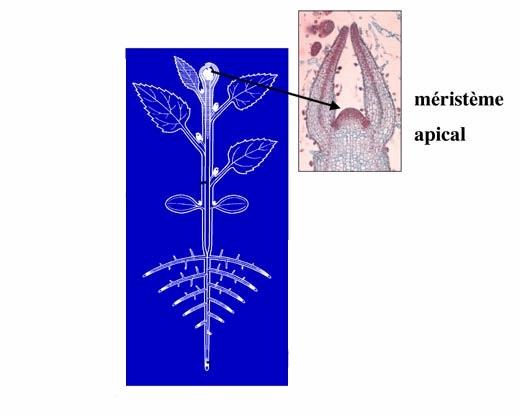
Les cellules à ce niveau se divisent sous l’action de l’Auxine puis se différencient pour devenir des cellules de feuilles, de vaisseaux conducteurs de sève, de tissu de soutient…
Ce sont également elles qui produisent le plus d’Auxine, cette Auxine va diffuser dans tout l’organisme végétal et cette différence de répartition va provoquer l’inhibition de la croissance des autres bourgeons.
Tant qu’ils sont trop proches de l’apex principal, ils ne grandissent pas, mais une fois qu’il est suffisamment éloigné, ils pourront grandir et former les branches secondaires, etc…
C’est pour cela que le Cannabis a un port comme les sapins.
Le pinçage vise à écraser et donc à détruire en grande partie les vaisseaux conducteurs de sève qui conduisent l’Auxine et donc, les autres apex peuvent se développer puisque le principal n’agit plus sur eux…
Le cas extrême étant bien sur le taillage, puisque là on supprime littéralement l’apex principal et donc son action inhibitrice sur les autres apex…
II.2. Le méristème racinaire :
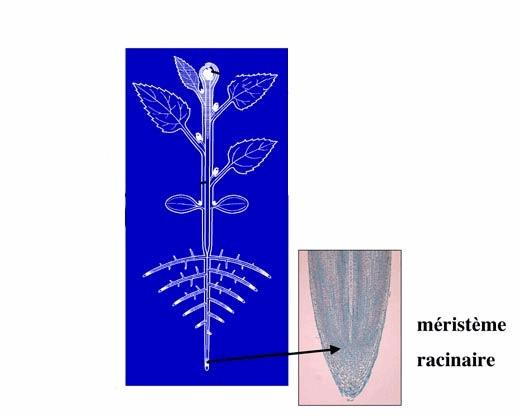
Il fonctionne selon le même principe mais la concentration en Auxine est différente…
Et puis aussi on ne peut pas aller tailler l’apex racinaire ^^ !
Donc on va pas trop s’éterniser sur les racines, c’est pas le plus important.
III. Les principales molécules intervenant dans le développement des plantes :
III.1. THE molécule, l’Auxine :
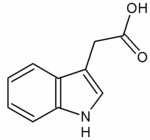
C’est une phytohormone (hormone végétale) qui est produite par les toutes les cellules, mais en petite quantité en général, sauf pour les cellules des apex…
A forte dose, elle augmente la vitesse de division cellulaire et donc la croissance de la plante par une activité plus importante des apex.
Mais elle intervient aussi énormément dans la différenciation des cellules de base :
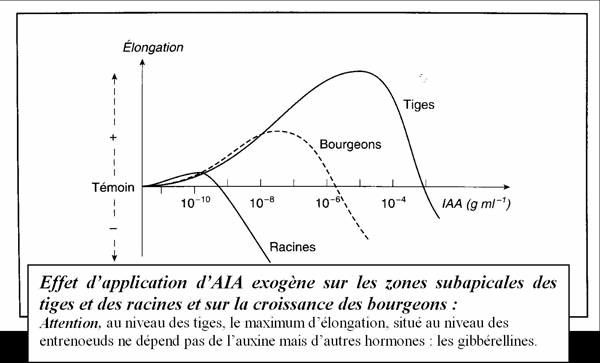
III.2. Les gibbérellines :
C’est en fait une famille extrêmement nombreuse de phytohormones qui interviennent dans le développement des tiges et des graines.
Pour tout savoir dessus :
Et donc pour résumer un peu tout ça : elles interviennent essentiellement dans la mérèse (allongement des cellules végétales) au niveau des tiges, et dans les graines dans la levée de certaines dormances (les dormances liées à l’acide abscissique).
III.3. L’acide abscissique :
Il est produit dans deux cas : le stress hydrique et la mise en place des graines.
Dans le premier cas, il est synthétisé par les racines en manque d’eau trop important et va agir au niveau des feuilles :
- fermeture des stomates au début, afin de limiter au maximum les pertes en eau
- mort des feuilles et chute de celles-ci pour limiter encore plus les pertes en eau par évapotranspiration (radical comme lutte !).
Dans le second cas, il est produit par la plante mère lorsque la graine est fabriquée après fécondation.
L’acide abscissique a pour rôle d’empêcher la germination avant qu’il ne se soit dégradé lentement jusqu’à une certaine concentration.
Cela prend plus ou moins longtemps selon des espèces végétales et ça permet à la graine de ne pas germer avant l’hiver (la plantule n’y survivrait pas).
La dormance permet donc dans ce cas d’éviter une germination précoce des graines…
IV. La photosynthèse :
C’est ce qui permet aux plantes de grandir, ce sont d’une part les ions absorbés au niveau des racines et d’autre part le CO² converti au niveau des cellules chlorophylliennes en matière organique (sucres, etc…).
IV.1. La conversion de l’énergie lumineuse:
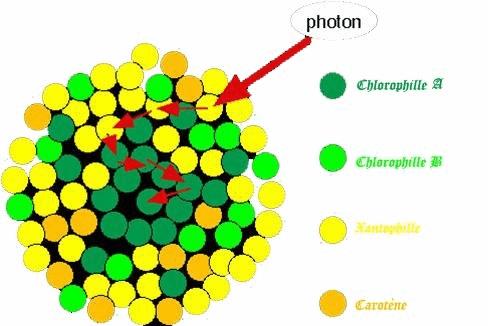
Cette opération se fait par l’intermédiaire des photosystèmes, molécules photosensibles contenues dans les membranes des cellules photosynthétiques au niveau de toutes les parties vertes de la plante (jeune tige, feuilles).
Cette énergie est ensuite utilisée pour mettre en place une énergie (grâce à un courant d’électrons) qui est utilisée pour produire de l’ATP, « monnaie énergétique des cellules».
IV.2. La fabrication de molécules organiques et fixation du CO² :
Cette étape se fait dans le cycle de Calvin :
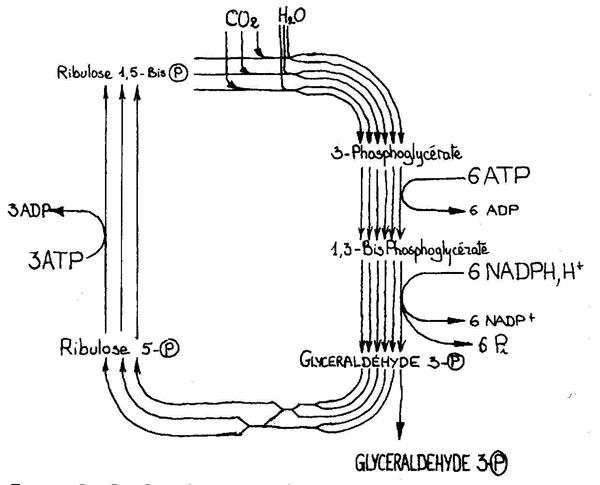
Le schéma peut être un peu difficile à comprendre mais bon en résumé, il y a fixation du CO² par une enzyme : la Rubisco, qui va transformer une molécule à cinq carbones en une molécule à six carbones et donc convertir le carbone atmosphérique en carbone organique et donc en sucres (pas le sucre de table, les oses en générale sont qualifiés de sucres).
En utilisant l’ATP produit lors de la phase précédente et d’autres molécules, les végétaux peuvent se rendre autonome pour ce qui est de l’approvisionnement en carbone (alors que les animaux doivent trouver leur carbone dans leur nourriture) et donc produire leur matière organique grâce au CO² de l’air, d’eau et de lumière.
C’est cette synthèse qui produit de l’O².
Mais en parallèle, toutes les cellules végétales respirent : c'est-à-dire qu’elles utilisent des dérivés du glucose, de l’O² et des molécules réductrices pour produire de l’ATP (encore lui, c’est la molécule la plus produite par les cellules) et rejettent du CO² dans l’air.
En journée, la production d’0² est plus importante que sa consommation et la consommation de CO² plus importante que sa production.
Donc le bilan en journée est : production d’O².
Mais la nuit, plus de production d’O² donc seule la respiration a lieu, d’où une production de CO² et une consommation d’O² durant la nuit.
V. l’absorption racinaire des nutriments :
Les nutriments captés dans le sol le sont sous forme d’ions :
NO3 -, NH4 +, Ca 2+, Mg 2+, Zn 2+, PO4 3-, K +, etc…
Tous ces ions sont absorbés par les racines au niveau des fragiles poils absorbants grâce à un échange de protons (ion H+).
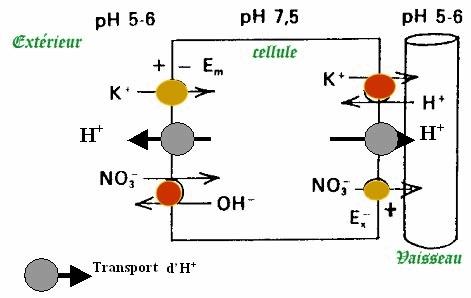
Le fait que le pH soit plus acide dans le milieu extérieur que dans les poils absorbants permet une entrée des ions qui libère de l’énergie lors du passage.
Cette énergie est utilisée pour faire entrer des ions qui ne peuvent entrer normalement car cette entrée nécessite de l’énergie.
Donc les ions H+ sont exportés via des pompes dépendantes à l’ATP (et oui, encore !) pour maintenir cette énergie, les nutriments sont absorbés grâce à la conversion énergétique décrite plus haut, et le même schéma se remet en place lors du passage des nutriments depuis les poils absorbants jusqu’aux cellules des vaisseaux qui conduiront les nutriments jusqu’aux feuilles pour qu’ils soient convertis en matière organique.
La différence de concentration de H+ entre les cellules et le sol (ou la solution Hydro) et la charge des ions donne la notion de potentiel électrochimique et le gradient électrochimique eu niveau de la membrane plasmique des poils absorbants permet en fonction des ions leur absorption ou non par la racine.
C'est pour ça qu'il y a un pH idéal pour l'absorption des nutriments: pH et nutrition par Jarhoïde
Il existe néanmoins des effets d'antognie entre certains ions qui peuvent entrainer des carences malgré tout apports d'engrais:
exemple pour le couple K+/Ca2+ ou K+/Mg2+. S'il y a trop de K+ dans le sol (ou la solution), sa forte mobilité et sa forte concentration empêche l'absorption des autres cations car ces derniers sont plus chargés et ont une sphère d'hydratation qui les rendent moins mobiles.
Ainsi, quelqu'un qui a la main un peu lourde sur le PK13-14 aura la surprise parfois avoir une carence en Mg ou Ca...
L’eau est absorbée au niveau de l’extrémité des racines (en majorité) et au niveau des poils absorbants grâce aux parois, puis par des aquaporines (protéines permettant un passage uniquement des molécules d’eau dans les cellules).
Plus il y a d'eau disponible dans le sol et plus l'absorption des nutriments est rapide et facile pour la plante.
Attention encore une fois à ne pas tomber dans l'extrême qui consiste à sur-arroser car les racines sont comme les humains: elles ne respirent pas sous l'eau !
VI. La gestion du potentiel hydrique :
Le potentiel hydrique correspond à la teneur en eau des tissus du végétal, s’il diminue trop, la plante se fane ou meurt si le déficit est trop important.
Bon, je suppose que tout le monde sait que les plantes captent l’eau au niveau des racines, mais le problème avec l’eau pour les végétaux (dans le nature) c’est qu’il y en a soit trop soit pas assez.
Si le sol est sec, la plante ne pourra plus puiser d’eau dans le sol et devra donc limiter le plus possible ses pertes en eau pour ne pas faner. Au contraire lorsqu’il pleut beaucoup, la plante se retrouve à la limite de la noyade et ses cellules sont envahies par l’eau, s’il n’y avait pas la paroi pour limiter la taille des cellules végétale, elles éclateraient à chaque grosse pluie ^^ !
Il est donc primordial de comprendre comment une plante absorbe de l’eau (voir partie absorption racinaire) et comment elle en perd.
VI.1. La notion d’évapotranspiration:
L’évapotranspiration, un nom barbare pour dire : transpiration par évaporation d’eau au niveau des feuilles.
C’est le facteur le plu important dans les pertes en eau.
Au niveau des feuilles il y a une cuticule (mince couche de lipides qui agit comme une bâche et qui retient l’eau au niveau de la feuille et empêche son évaporation), plus ou moins épaisse selon les espèces, qui empêche en partie l’évapotranspiration.
Mais c’est au niveau des stomates qu’elle est la plus importante.
Les stomates sont des cavités formées par un complexe de deux cellules de gardes, évoluant d’une position ouvert à fermé sous l’effet de l’acide abscicique.
L'orifice formé par la baisse de turgescence s'appelle l'ostiole et est le résultat de l'adaptation à la vie terrestre des végétaux. Il permet les échanges gazeux et surtout le maintient d'un statut hydrique positif, malgré le phénomène de plasmolyse.
Il sont très majoritairement situés sous les feuilles (pour le cannabis) en corrélation avec une cuticule présente sur la face supérieure limitant la déshydratation :
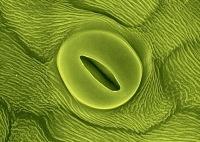
En dessous du stomate se trouve une cavité : la chambre sous-stomatique, au niveau de laquelle se font les échanges d’air entre les cellules végétales :
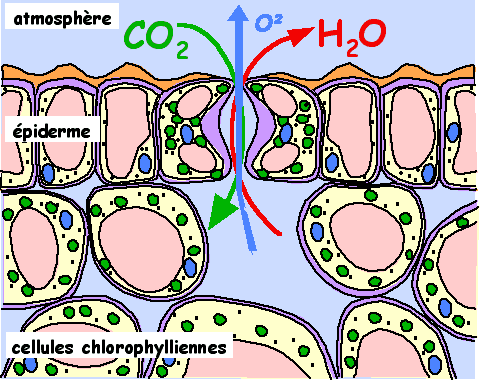
Pour vous donner une idée de l’importance de l’évapotranspiration, un plant de maïs qui absorbe 100L d’eau en rejette 90L par évapotranspiration… Donc quand on voit que l’irrigation du maïs assèche les nappes phréatiques et les cours d’eau, pollue, etc… dans certaines régions, on se dit qu’il serait temps de changer de culture ^^ !
NB: pour plus de précisions sur un concept plus précis: Les Osmoticums par Exo
VI.2. Les contrôles des pertes en eau:
Ils passent essentiellement par un contrôle de l’ouverture et de la fermeture des stomates, puisque c’est à ce niveau là que les plantes perdent le plus d’eau.
Les facteurs suivant favorisent l’évapotranspiration :
-environnement sec (air chaud et sec ou froid et sec, terre sèche)
-chaleur importante
-vent
-manque de CO²
Lorsque ces facteurs sont présents, des molécules sont sécrétées par la plante (comme l’acide abscissique au niveau des racines lorsque le sol est trop sec) et entrainent la fermeture des stomates.
Au contraire, certaines longueurs d’ondes lumineuses comme le bleue et le rouge lointain (far red en english) tendrait plutôt à les ouvrir.
Dans tous les cas, les plantes ont plutôt tendance à ouvrir leurs stomates en début de journée, et durant la nuit car l’air est plus frai et l’humidité est plus importante.
C’est pour ça qu’une forte humidité en croissance est importante : une plantoune qui est en croissance et qui, dès qu’elle ouvre ses stomates se déshydrate très rapidement, va les garder fermées.
Donc elle absorbera moins de CO², donc elle grandira moins vite et aura des feuilles qui risquent de tomber ou d’être mal-formées…
Donc pensez bien à conserver une humidité d’au moins 60% pour votre croissance !
Et ne pas mettre un ventilo puissant en brassage de l’air, ça dessèche nos petites !
VII. Les dormances:
VII.1. Le principe de dormance:
Dormance : adjectif utilisé pour parler d’une graine ou d’un bourgeon qui ne grandit pas et dont l’activité métabolique est au ralentit (presque à l’arrêt en fait).
On distingue la dormance de la simple vie ralentie car un bourgeon ou une graine en vie ralentie va germer dès que les conditions favorables apparaissent alors qu’une graine encore sous l’effet de dormances ne germera pas, même si les conditions sont favorables.
Plusieurs facteurs peuvent intervenir dans le phénomène de dormance, nous allons développer un petit peu les principaux facteurs…
VII.2. Les dormances tégumentaires:
Les téguments de la graine sont les enveloppes plus ou moins rigides (la coquille qui ressemble à une carapace de tortue chez la graine de Kaya, ce sont les téguments externes de la graine !), ils jouent un rôle protecteurs le temps que la graine passe l’hiver, ils les protègent des pertes d’eau en partie, et empêchent le germe de sortir de la graine car ils sont très solides au moment de la libération de la graine.
Hors, la graine est libérée en générale en Octobre, juste avant la saison froide qu’est l’hiver.
Et une graine qui se mettrait à germer fin Octobre ne pourrait pas vivre très longtemps avec la chute brutale des températures et le manque croissant de soleil !
Donc la solidité des téguments externes (coquille) protège la graine d’une germination précoce qui lui serait fatale !
Cette dormance est levée durant l’hiver par les micro-organismes du sol et par l’humus (acides organiques que l’on trouve dans la partie superficielle du sol, là où tombent les graines). Les micro-organismes vont attaquer en partie la surface de la graine et ainsi la fragiliser, alors que l’humus va encore fragiliser les téguments à cause de son acidité.
A la fin de l’hiver, si les autres dormances ont aussi été levées, le retour des conditions favorable permet à la graine de germer…
VII.3. Les dormances embryonnaires:
Bon, ce sont les plus complexes en fait au niveau du mécanisme alors on fera dans le simple pour ce premier jet, si jamais vous avez besoin/envie d’une version plus précise, je verrai un peu plus tard si je peux compléter tout ça…
Alors en gros, au moment de la formation de la graine, la plante produit de l’ABA (ABscissique Acid) en quantité importante et cet ABA est concentré au niveau de l’embryon dans la graine.
On constate que même dans des conditions favorables, une graine contenant trop d’ABA ne germe pas, c’est donc un des facteurs qui gardent la graine en dormance.
Or l’ABA est une molécule organique, elle se détruit donc au bout d’un certain temps, qui comme par hasard correspond à 1 mois ou 2 ce qui amène notre petite graine en Décembre.
Mais si elle n’est plus inhibée en Décembre, elle risque de germer ?!
Et non car en Décembre il fait en général très froid ^^, on peut germer, mais on n’a pas les conditions favorables donc on ne germe pas !
Dans l’étude du coléoptile de blé (du grain de blé, mais coléoptile ça pète plus ^^), on a montré que si l’on augmentait la concentration en certaines gibbérellines (et oui, encore et toujours les gibbérellines), l’action inhibitrice de l’ABA était fortement diminuée :
A concentration équivalente, les Gibb. sont dix fois plus influentes que l’ABA, donc une très faible dose suffit à lever la dormance liée à l’ABA.
Dans la nature, ces Gibb. sont produites par l’embryon lorsque les conditions favorables reviennent.
La levée de dormance est ici due à un équilibre entre l’action de deux hormones principales, ce qui fait la complexité du mécanisme…
Ici encore, il faut que les autres dormances ait été levées, sinon la graine reste en dormance.
VII.4. Les dormances psychrolabiles :
C’est le dernier type de dormance courante, elle agit sur l’embryon également mais je ne pourrai pas détailler trop le mécanisme car je ne le connais pas, mais il doit être comparable à celui des dormances chimiques embryonnaires.
Psychro=froid, c’est une dormance levée par un passage au froid…
En gros : tant que la graine n’a pas passé une période suffisamment importante dans le froid, elle ne peut germer.
En horticulture, on utilise la technique de « stratification » pour lever la dormance des graines :
On fait une strate de tourbe (acide) une strate de graines, tourbe, graine….
Et l’on arrose le tout en conservant ces piles dans une chambre froide avec une température proche de 1-2°C.
La levée de dormance décrite dans les guides : congélateur, frigo, endroit noir et sec permet de lever la dormance psychrolabile par le passage au congélo et la dormance à l’ABA par le passage en endroit sec (la sècheresse augment la vitesse de disparition de l’ABA).
Alors bien sûr, il n’est pas nécessaire de laisser vos graines issues de cross persos 2 mois au congélateur, mais en suivant le guide de levée de dormance, ça devrait aller…
Ainsi, les végétaux grandissent grâce à la division des cellules des méristèmes situés au niveau des apex, ces cellules se différencient sous l’action de phytohormones très nombreuses et dont les concentrations déterminent le devenir de ces cellules.
Ils captent les nutriments et l’eau contenue dans le sol grâce aux précieux poils absorbants et à l’existence d’une différence de pH entre les cellules et le milieu extérieur pour ensuite les apporter jusqu’à toutes les autres cellules via les vaisseaux conducteurs de sève.
Pour ce qui est de la reproduction et des graines, il existe des guides qui en parlent…
A bientôt !
PS : s’il y a des erreurs selon vous, ou des points encore trop sombres, faites m’en part et je tenterai de modifier tout ça… 
PS2: topic sur la micropropagation par Exo: ICI
[edit]MAJ le 21.06.09[/edit]
v/c Dad-
-
 2
2
-
 1
1