


Résistance aux pathogènes - S.A.R
I- Introduction
Au cours de leur vie, nos plantes peuvent être confrontées à différents agents pathogénes ou blessures mécaniques (herbivores).
On appelle agents pathogénes :
* les bactéries,
* les nématodes,
* les mycoses,
* les virus.
A chaque type d'agression, la plante y répondra de façon spécifique.
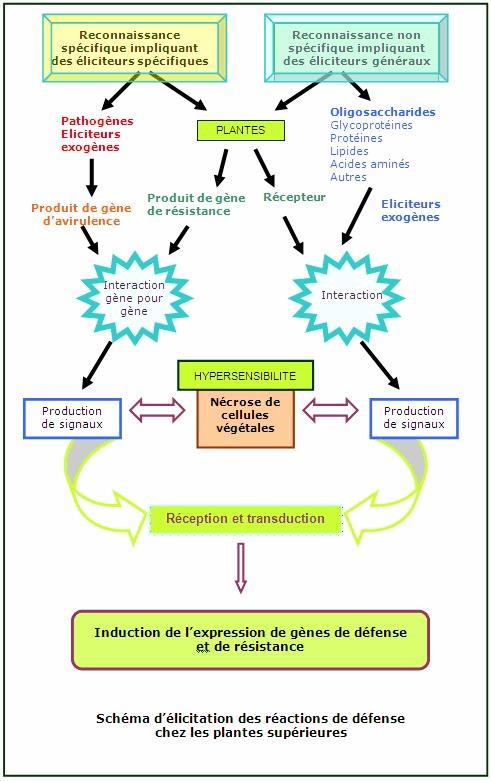
II- Processus de reconnaissance
Deux cas peuvent se produire, soit la plante ne reconnait pas l'agresseur et là malheureusement sa mort est inéluctable soit elle arrive à reconnaitre l'agent pathogéne et là va se mettre en place la résistance de la plante face à l'agression.
Cette reconnaissance pour être efficace doit se produire rapidement.
Pour cela, il faut que la plante posséde un nombre suffisant de protéïnes sentinelles présentes au niveau de la cellule végétale ainsi qu'avoir la possibilité de reconnaître la présence de l'agent infectieux à de trés faible concentration.
Pour les attaques virales, fongiques, bactériennes et de nématodes, ces reconnaissances se font en dehors de la cellule végétale, avant qu'ils n'aient le temps de pénètrer à l'interieur de celles-ci.
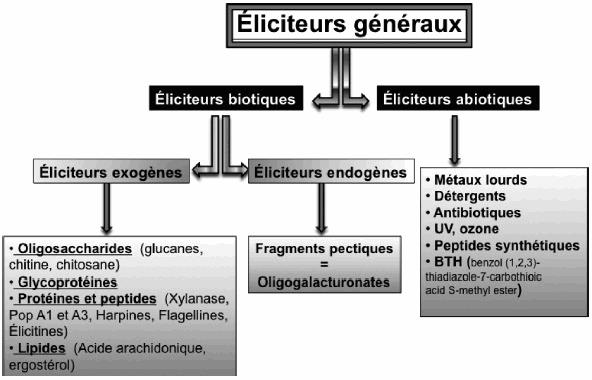
On parlera de pathogéne avirulent si la plante arrive à le reconnaître ou encore éliciteur et dans le cas où l'agent infectieux n'est pas reconnu on l'appellera pathogène virulent.
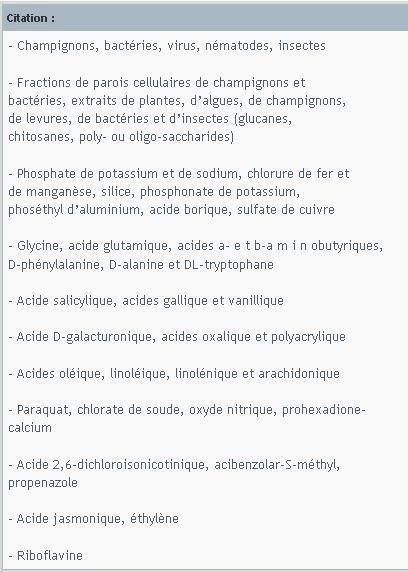
Liste des éliciteurs :
III- La réponse Hypersensitive(HR)
On parle de réponse hypersensitive car dés que l'agent pathogéne infecte la cellule hôte saine, celle-ci meurt instantanement. C'est la premiére réponse qu'a la plante face à son agresseur.
Afin de se défendre, la plante va mettre en oeuvre une réponse adaptée soit :
- Le renforcement de la paroi membranaire de la cellule agressée est la réaction de défense qui est mise en oeuvre le plus rapidement. C'est aussi une des réactions les plus communes chez les plantes. Elle se traduit par la modification des parois des cellules de la plante au niveau du site d'inoculation de l'agent pathogène.
Des dépots de cellulose, de callose et de subérine au niveau de la parois cellulaire vont la renforcer et la solidifier.
- Il va se produire une modification chimique de celle-ci ( production de Phénols oxydés, mélanines, calcium, silice, soufre, lignine), par exemple les précurseurs de la lignine sont toxiques pour les micro-organismes.
- une modification des molécules de la parois (quantité, type, formation de liaisons oxydatives) formant ainsi une sorte de mur physique.
- une nécrose (mort) des cellules infectées. La plante va en fait sacrifier quelques cellules afin de ralentir ou stopper l'infection.
Ces foyers nécrotiques sont le résultat d'une production par la plante de péroxyde d'hydrogéne (H2O2), on appelera alors les processus une péroxydation et d'oxygène (O2).
On observe notamment un influx d'ions Ca2+ et un efflux d'ions K+, une dépolarisation, une vacuolisation ( formation de vésicules liées à la membrane du cytoplasme), la mise en œuvre d'endonucléases (Une endonucléase est une nucléase, qui coupe un acide nucléique en fragments plus courts, On parlera de digestion de l'ADN de l'agent infectieux).
Exemple de foyers nécrotiques suite à une anthracnose :

- La synthèse d'intermédiaires oxygénés réactifs ou Reactive Oxygen Intermediates ou encore R.O.I.
Les ROI sont des composés cytotoxiques. Une régulation fine est donc vitale. Les mécanismes de régulation font intervenir des réactions de phosphorilation/déphosphorilation et mettent en jeu l'ion Ca2+.
la HR intervient tout d'abord dans la mise en place des mécanismes des interactions entre pathogènes biotrophes (comme par exemple certains champignons) et la plante. La mort immédiate de la cellule infectée empêche en effet l'établissement d'une relation effective entre les deux intervenants. La croissance du pathogène est donc inhibée par la suppression de ses moyens de nutrition.
Mais les pathogènes nécrotrophes (certains champignons et bactéries) peuvent aussi déclencher une réaction hypersensitive. En effet, la mort des cellules infectées empêche aussi leur développement.
Enfin, la nécrose des cellules autour du site d'infection des virus empêche son développement dans tout l'hôte.
La mort programmée des cellules de la plante autour du site infecté ne détermine pas le sort du pathogène.
En conclusion de ce chapitre, nous pourrons dire que tous ces évènements permettent donc de créer un environnement d'inhibition pour le pathogène : soit la croissance du pathogène est réduite, soit le pathogène est affamé et/ou empoisonné, soit il est arrêté par une barrière physique.
Dans d'autres cas, la HR semble plutôt être une conséquence de l'attaque du pathogène : c'est donc le pathogène lui-même qui provoque la mort de la cellule qu'il infecte.
IV- La Réponse Acquise Systémique (SAR)
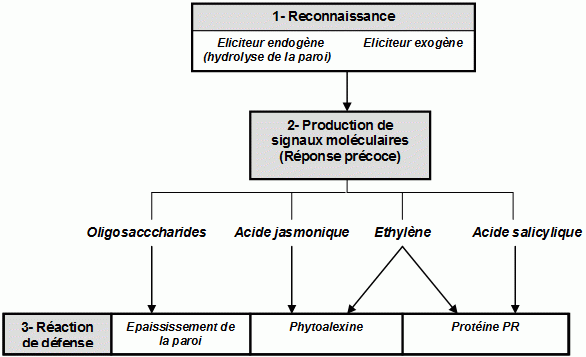
La réponse systémique acquise (SAR) se met en place à la suite de la HR. Elle est classée dans le groupe des réponses systémiques.
Les réponses systémiques peuvent être assimilées à l'établissement d'une immunité de la plante. L'infection par un pathogène va en effet entraîner la mise en place d'une immunité pour les infections futures dans l'ensemble de la plante.
De nombreuses réponses systémiques inductibles ont été découvertes durant les 10 dernières années. Certaines sont passives, et d'autres actives.
Une fois activée, la SAR procure une protection de longue durée contre toute attaque ultérieure par des micro-organismes pathogènes.
C'est une infection au niveau local qui entraîne la mise en place de cette réponse globale dans toute la plante.
Tous les exemples de mort de cellules ne provoquent cependant pas l'activation de la SAR. Certains seulement des phénomènes de nécrose pourraient provoquer la SAR.
Il existe divers signaux systémiques qui vont donc déclencher la S.A.R, ces messagers sont l'acide salicylique, les proteïnes PR, l'acide jasmonique,Les phytoalexines et enfin l'éthylène . En effet, ces molécules semblent jouer un rôle trés important dans les processus de mise en place de la SAR.
V- L'acide salicylique (SA) :
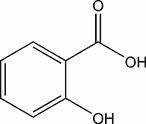
Composé phénolique issu de la voie des phénylpropanoïdes qui active la synthèse des protéines de défense. C'est un acide carboxylique incolore et cristallin. Il Participe à la mort cellulaire lors de la réaction d’hypersensibilité et semble induire la résistance systémique acquise. Cette molécule joue un rôle essentiel dans l’alerte des cellules.
On a observé qu'après l'infection par des pathogènes, la concentration d'acide salicylique devient élevée dans toutes les cellules des feuilles non infectées de la plante. Cette accumulation de SA paraît nécessaire à l'établissement de la SAR.
Cette molécule joue donc un rôle clé dans la transduction (Transformation d'une grandeur physique ou d'une énergie en une autre qui est fonction de la première) du signal systémique menant à l'établissement de la SAR. Mais la nature de ce second signal reste encore inconnue de nos jours.
Il peut agir comme un signal hormonal pouvant déclencher, dans certains cas, une thermogénèse végétale déclenchée par la sécrétion d'une molécule, le calorigène.
Les protéïnes PR:
Les protéines PR sont des protéines de stress végétale, c'est-à-dire protéines reliées à la pathogénèse.
Tout un panel de protéines ont été identifiés en relation avec l'établissement de la SAR. Ce sont les pathogenesis-related proteins, ou protéines PR.
Aucune de ces protéines n'existe naturellement dans les plantes, elles sont toutes synthétisées suite à une attaque par un pathogène.
La production de PR est induite par diverses molécules chimiques, incluant des hormones comme l'éthylène. Dans ce cadre, la synthèse de PR peut se faire pour divers types de réaction de stress, y compris la SAR.
De plus, l'expression des gènes PR varie selon les plantes. Par exemple, on retrouve les 5 types chez le Tabac, mais seulement PR2 et PR5 chez les Arabidopsis.
Les PR s'accumulent ensuite principalement dans l'espace intercellulaire du tissu des plantes. Les PR1 et PR4 ont une activité anti-fongique, les PR2 et PR3 auraient une action enzymatique. L'expression des protéines PR a donc un rôle direct sur le ralentissement de la croissance des pathogènes, et un rôle indirect par la destruction des parois des cellules de ce même pathogène. Les résidus de cette dégradation sont en effet souvent des signaux permettant à la plante d'identifier le pathogène.
VII- L'acide jasmonique :
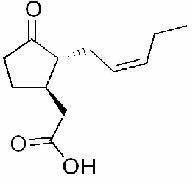
Dérivés de la famille des acides gras octadécanoïques synthétisés à partir de l’acide linoléique. Il contribue à l’activation des protéines de défense. Cette molécule clé intervient dans la propagation du signal au niveau de la cellule végétale.
L’acide jasmonique est un messager secondaire dans la cascade de signalisation de la réponse de la plante à un stress.
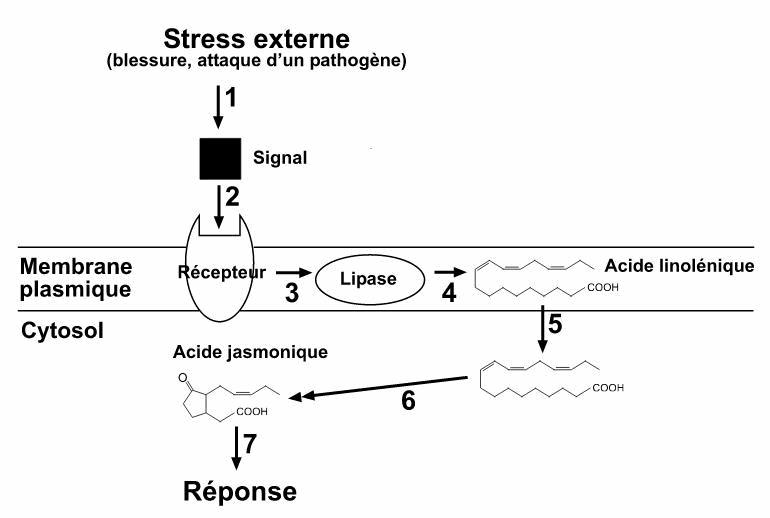
Voie de signalisation par l’acide jasmonique. Un signal extérieur est transformé en signal chimique (1), un récepteur membranaire reçoit le signal (2), et active rapidement une lipase (3), qui libère de l’acide linolénique (4 et 5). Ce dernier est convertit en acide jasmonique par cinq réactions enzymatiques (6). L’acide jasmonique peut induire différents types de réponses/défenses(7).
VII- Les phytoalexines:
Les phytoalexines (Du grec phyton = plante , et alexein = protéger) sont des molécules produites par la plante suite à une infection ou un stress. Elles jouent un rôle de pesticide naturel et jouent un rôle d'inhibiteur de l'agent infectieux. Elles sont, en général, absentes des plantes saines (Keen, 1971; Darvill et Albersheim,1984). Il s’agit de composés de stress pouvant être synthétisés et s’accumuler en réponse à des contaminations (virus,bactéries, champignons),des attaques par des insectes ou des nématodes, mais aussi à de nombreux stimuli abiotiques(éthylène, froid, UV, fongicides, sels métalliques, détergents, stress dus à l’environnement) ou à des élicitations biotiques (composantes de parois bactériennes, fongiques ou végétales,enzymes microbiennes, etc...
Chaque espèce végètale possède ses propres phytoalexines. Le terme phytoalexine a été créé par Müller et Borger (1940) pour désigner des substances antifongiques.
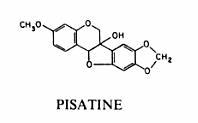
Les chercheurs ont observé la mort rapide des cellules végétales, au niveau du point de pénétration du parasite, accompagnée d’une réaction de défense des cellules environnantes. Les cellules ayant ainsi réagi sont immunisées contre de nouvelles attaques du parasite. Cette réaction de défense reste très localisée et n’entraîne jamais une prémunition de l’individu entier. Müller et Borger lient cette résistance à l’activation ou à la formation dans les cellules de l’hôte d’une substance de défense non spécifique ou phytoalexine, occasionnant la paralysie ou la mort du parasite. Selon ces auteurs, la résistance, bien que transmise génétiquement, n’est pas constitutive en ce sens qu’elle ne s’exprime qu’après le contact avec l’agent pathogène. Plus de 20 ans après les travaux de Müller et Borger, les premières phytoalexines la pisatine et la phaséolline ont été isolées puis caractérisées (Cruickshank et Perrin, 1960, 1963; Perrin et Bottomley, 1962). Une définition de référence a été énoncée par un groupe de chercheurs (Paxton, 1981) : Les phytoalexines sont des composés antimicrobiens.
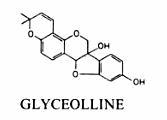
Lors d’une attaque parasitaire, la réponse d’une plante résistante peut être extrêmement rapide, puisque Hahn et al. (1985) détectent les premières traces de glycéolline dans des racines de soja 2 h après la contamination.
L’accumulation est très retardée et/ou moins intense chez les plantes sensibles à un parasite et ne présentant pas (ou peu) de réaction hypersensible à ce dernier.
Molécule de la pisatine du pois :
Molécule de la glycéolline du soja:
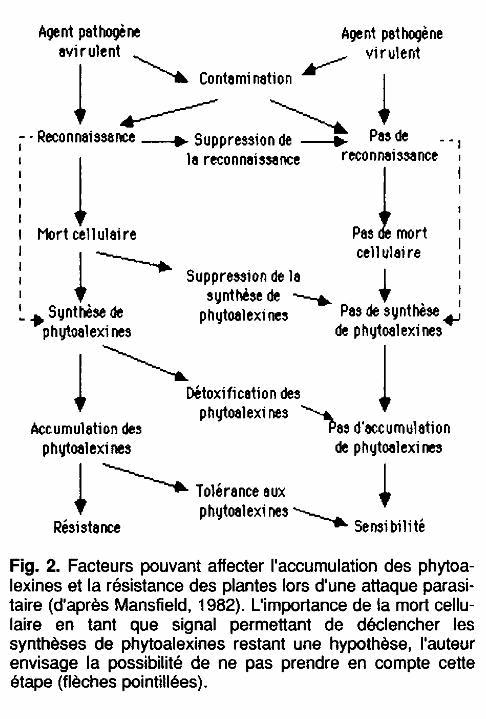
Récapitulatif schématique de la production des phytoalexines:
VIII- L'éthylène :
https://wiki.cannawee....A9thyl.C3.A8ne
IX- Conclusion :
Nous avons pu voir tout au long de cet exposé, la complexité et la multiplicité des phénomènes de défense de la plante.
Transposé au cannabis, toutes ces voies de résistance peuvent être des plus interessantes et prometteuses afin d'en améliorer leur rendement.
X- Bibliographie :
Cours perso.
Internet.
-
 1
1