


La Sexualité du Cannabis
Le cannabis est une plante généralement dioïque [ du grec "dis ", (deux fois), et "oikos ", (maison) ]
c'est-à-dire plante ayant des fleurs mâles (à étamines) sur un pied et des fleurs femelles (à pistils) sur un autre pied.
Le sexage des plantes de cannabis est important car seules les fleurs femelles portent une couverture intéressante de trichomes glandulaires.
Exemples :
Une femelle :


Un mâle :


edit avril 2017
magnifique exemple de mâle par animalxxx

La déclaration des sexes :
Le cannabis fleurit lorsque la photopériode atteint un seuil critique différent selon les variétés.
Une durée de nuit longue est nécessaire pour enclencher la production de fleurs fertiles.
Autrement, il ne produira que des primordiae indifférenciées.
Pour en savoir davantage : [ Traduction ] Article de Ed Rosenthal
Durant la phase de croissance sous une durée de nuit courte, il est possible d’observer l’apparition des primordiae ou premiers pistils à partir du 5ème nœud. Ces pré-fleurs servent d’indicateurs précoces du sexe de la plante.




Ce stade est atteint lorsque la plante est suffisamment mature. Les signes avant-coureurs les plus fréquemment observés sont :
-le passage d’une phyllotaxie décussée (opposée) à alternée (décalée)
-des modifications de la tige
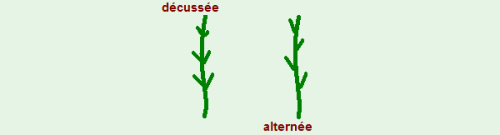
Il est possible de procéder à un sexage précoce à partir d’un certain nombre de caractères végétatifs
cependant cela reste extrêmement difficile pour un débutant (Lacombe, 1987).
nota bene : - De l'utilisation du terme pré-floraison :
Il est essentiel d'éviter ce terme car il n'apporte que confusion.
Défini comme la période allant de l'apparition des pré-fleurs à la floraison (apparition des fleurs "vraies"),
il est abusif de l'employer car :
- En culture intérieur, l'apparition des pré-fleurs peut se faire sous un cycle de croissance
et on ne va pas parler de pré-floraison alors que nous sommes sous un cycle 18/6 (croissance ).
- En culture intérieure, les pré-fleurs peuvent apparaître bien après le passage en 12/12.
Alors comment nommer cette période où les pré-fleurs sont absentes mais sous un cycle 12/12 (floraison ) ?
Il faut lui préférer le terme de stretch. Le seul cas où ce terme est approprié, c'est lors de départ de graines en extérieur
(et encore ... )
La diagnose des sexes :
Premier point, ne pas confondre les stipules avec des primordiae.
Les stipules sont des petites pièces foliacées, qu'on trouve à la base du pétiole à l'endroit où il s'insère dans la tige.
Et elles sont souvent confondues avec des pré-fleurs.


Les pièces florales visibles de chaque coté des bourgeons axillaires (dans le cercle rouge) sont bien des stipules et en aucun cas ne les confondre avec des primordiae.
Les pré-fleurs mâles :
Elles se caractérisent par la présence de :
-5 pétales (longueur environ 5mm) forment le calice, de couleur indifféremment jaune, blanche ou verte
-5 étamines (d’environ 5mm ) formées d’anthères (sacs à pollen)


Elles apparaissent soit en croissance sous un cycle avec une période de nuit courte (entre 4 à 12 semaines selon les variétés, les sativas mettant plus de temps à atteindre la maturité sexuelle), soit sous un cycle avec une période de nuit longue (floraison ) au bout de 1 à 3 semaines ....
En général, les pré-fleurs mâles apparaissent plus rapidement que les pré-fleurs femelles.
Les pré-fleurs femelles :
Elles se présentent au départ sous la forme de deux longs pistils blancs, jaunes ou roses qui émergent de la fente d’un calice.
Le calice mesure de 2 à 6mm et contient l’ovule qui va donner la future graine.


Si vous êtes incertain de votre diagnose, il vous suffit d'attendre quelques jours que les organes soient mieux formés et caractéristiques.
A ce stade, vous avez plusieurs semaines avant que les inflorescences ne se forment réellement.
Vous pouvez cependant vous rendre sur le [ Topic Unique ] Sexage : Mâle ou Femelle ?
Les inflorescences adultes males :
Une fois les grappes de fleurs parfaitement formées comme sur les photos [plusieurs semaines de floraison],
Il s'agit d'une question de jours avant qu'elles ne libèrent le pollen.


Les inflorescences adultes femelles :
Ce sont celles qui nous intéressent et particulièrement si elles sont sensemilia,
c'est pourquoi il est important de les séparer rapidement des mâles et des intersexués



* Visitez des JDC 
Le cas particulier des intersexes :
Le cannabis est une plante possédant un grand polymorphisme dans son expression sexuelle.
Communément appelées « hermaphrodisme », il est plus juste de désigner ces anomalies sous le nom d’ inter-sexes
(Borodina & Migal , 1987). En effet, l’hermaphrodisme végétal exprime plutôt la présence sur une même fleur d’organes males (étamines ) et femelles (pistils ).
La monoécie :
Il s’agit d’un cas particulier d’inter-sexe dans lequel les plantes portent des fleurs mâles et femelles parfaitement formées.
Elles apparaissent le plus souvent lors des premiers stades de la floraison . Et les plantes incriminées doivent être éliminées le plus rapidement possible sauf si le phénomène est très limité (à peine quelques fleurs) en retirant les fleurs mâles.
Mais cela reste un pis-aller et un risque important

{kind=link}
On aperçoit ici la coexistence de fleurs femelles (pistillées) au sommet de l'inflorescence et de fleurs mâles (staminées ) à la base.
Les fleurs tératologiques :
Ce sont des fleurs dans lesquelles coexistent à des degrés divers des pièces florales mâles et femelles.
Elles ont tendance à apparaître plus tardivement durant la floraison et sont plus difficiles à déceler.



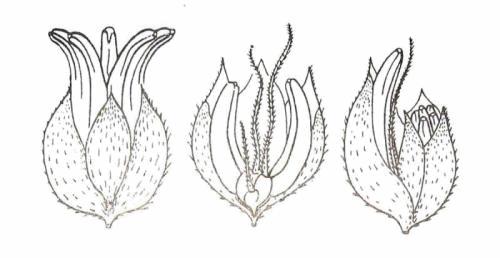
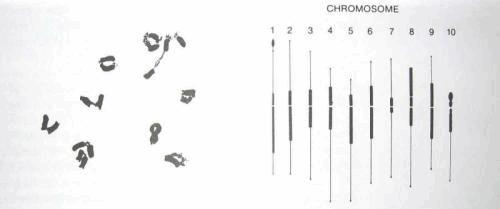
La détermination du sexe
Le cannabis est un organisme généralement diploïde (2n chromosomes ) avec n égal à 10 (Menzel ,1964) .
Parmi ces 20 chromosomes, 9 paires d’autosomes et 1 paire d’ hétérosomes.
Cette paire de chromosomes sexuels est constituée soit :
- de deux chromosomes X et le plant sera génétiquement femelle
- d’un chromosome X et Y, génétiquement mâle
Le plant de cannabis mâle est une plante hétérogamétique, c'est-à-dire que les hétérosomes sont différents. Cependant, la détermination sexuelle serait plutôt basée sur l’équilibre X-autosome que sur un mécanisme Y actif
(Westgaard, 1958; Grant et al., 1994).
De nombreuses études récentes ont mis en évidence l’existence de marqueurs sexuels spécifiques ( Mandolino et al.,1998, 1999) tendant à prouver l’origine génétique de la détermination sexuelle. Quant aux inter-sexes, ils ont eux aussi une origine génétique comme le prouve la capacité de certaines variétés à subir une conversion sexuelle aisément (Fibranova cv., landraces Thailandaises ) ou au contraire à y être très résistantes (ecotype italien Carmagnola).
Les origines de ce caractère seraient :
-la polyploïdie (Warmke et Zhatov)
-l’existence d’un ou plusieurs allèles directement responsables de l'intersexualité par le biais "d'un mécanisme polyfonctionnel de détermination sexuelle [chez le cannabis intersexué] contrôlé par l'interaction d'allèles appartenant aux chromosomes sexuels et d'allèles appartenant aux "autosomes" (Migal).
L'extension de l'expression d'un phénotype intersexué est quant à elle en partie dépendante des conditions du milieu (stress divers et variés, pollution lumineuse, ... ).
Le cannabis est une espèce possédant une grande plasticité dans l'expression de son phénotype sexuel ce qui a quelques conséquences non négligeables.
Conséquences et applications pratiques :
L'application évidente est bien entendu la modification du ratio mâle / femelle qui est normalement de 50/50.
A ce sujet, ces statistiques sont établies sur un nombre extrêmement élevé de graines. Un échantillonnage aussi réduit que ceux que l'on utilise est donc forcément biaisé et l'occurrence d'un paquet avec zéro femelle est hélas non nulle.
L'exploitation de ce phénomène est ancienne et se base sur le croisement de plantes monoïques et dioïques femelles.
(McPhee, von Sengbusch and Hoffman, 1943 ). Le résultat de tels croisements permet d'obtenir une population F1 dont la répartition du phénotype sexuel est :
- 90% de femelles
- 3-5% de monoïques
- 4-5% de mâles
Ce type de population constituée quasi en totalité d'individus à graines est dénommée unisexuée. Elle possède un intérêt certain dans la production de graines à fins industrielles.
Cependant, pour l'obtention de sinsemilla, il s'agit d'une véritable plaie car cela demande une attention constante afin d'éliminer les individus monoéciques sans compter que ce caractère peut resurgir de façon impromptue...
Cette méthode ne permet donc pas d'obtenir des populations de femelles stables étant donné la présence de facteurs génétiques responsables d'inter-sexes au sein de la population F1 obtenue.
L'alternative la plus intéressante serait donc de réussir à faire produire à des femelles non-porteuses de ces allèles des fleurs mâles afin d'obtenir une population F1 de femelles stables. Galoch (1980 ) a démontré l'importance de l'induction de gènes responsables de la différenciation en femelle par l'auxine. Ces travaux ont ouvert la voie à diverses méthodes d'induction de fleurs mâles sur des individus dioïques femelles notamment par :
- le nitrate d'argent et le thiosulfate d'argent anionique ( Mohan, 1982 )
- le GA3
Les populations F1 obtenues sont stables (pas d'apparition d'inter-sexes) à la condition évidemment que les parents ne possèdent pas les allèles responsables de l'inter-sexe. Ce qui est loin d'être facile à déterminer à l'heure actuelle
Une autre méthode est de conserver le pollen pour une assez longue durée (supérieure à deux semaines ) ce qui a pour effet de sélectionner un pollen qui donnera une population à majorité femelle (laskowska, 1961).
Conclusion :
En simplifiant, on peut dire que la détermination du sexe est génétique. Cependant, dès que l'on aborde la question des inter-sexes, cela se complique car il s'agit d'un mécanisme polyfonctionnel :
- un, et plus probablement, plusieurs allèles sont impliqués
- ces allèles sont répartis dans les autosomes et les hétérosomes
- leur activation est sujette aux conditions du milieu
De plus, les techniques d'inbreeding actuelles et la diminution de l'hétérosis dans les populations
ne sont pas étrangères à la fréquence des phénomènes d'intersexualité.
Pour aller plus loin => Génétique et Breeding par Cr4b
Bibliographie :
- Borodina E. I. and N. D. Migal 1987. Flower teratology in intersexual hemp plants. Soviet Journal of Developmental Biology 17(4): 262-269.
- Galoch, E., 1980. The hormonal control of sex differentiation in dioecious plants of hemp (Cannabis sativa). Acta Physiol Plant vol II (n.1): 31–39.
- Grant, S., A. Houben, B. Vyskot, J. Siroky,W.H. Pan, J. Macas & H. Saedler, 1994. Genetics of sex determination in flowering plants. Dev Genet 15: 214–230.
- Lacombe, J.-P., 1980. Discrimination des sexes en fonction de caractères végétatifs précoces chez le Chanvre dioïque (Cannabis sativa L.). Physiol Veg 18: 419–430.
- Laskowska, R.: Influence of the age of pollen and stigmas on sex determination in hemp. Nature, 1961, 192, No. 4798, p 147-148
- Mandolino, G., A. Carboni, S. Forapani & P. Ranalli, 1998. DNA markers associated with sex phenotype in hemp (Cannabis sativa L.). In: Proc Bast Fibrous Plants Today and Tomorrow, St Petersburg, September 28–30, pp. 197–201.
- Mandolino, G., A. Carboni, S. Forapani, V. Faeti & P. Ranalli, 1999.Identification of DNA markers linked to the male sex in dioecious hemp (Cannabis sativa L.).Theor Appl Genet 98: 86–92.
- Mandolino, G., A. Carboni, M. Bagatta, V.M.C. Moliterni & P. Ranalli, 2002. Occurrence and frequency of putatively Y chromosome linked DNA markers in Cannabis sativa L. Euphytica 126: 211–216.
- Mandolino, G. & P. Ranalli, 2002. The applications of molecular markers in genetics and breeding of hemp. J Ind Hemp 7: 7–24.
- Menzel, Margaret Y (1964). "Meiotic Chromosomes of Monoecious Kentucky Hemp (Cannabis sativa)". Bulletin of the Torrey Botanical Club 91 (3): 193-205.
- Mohan Ram, H.Y. & R. Sett, 1982a. Modification of growth and sex expression in Cannabis sativa by aminoethoxyvinylglycine and etephon. Z Planzenphysiol Bd 105: 165–172.
- Mohan Ram, H.Y.&R. Sett, 1982b. Induction of fertile male flowers in genetically female Cannabis sativa plants by silver nitrate and silver thiosulphate anionic complex. Theor Appl Genet 62: 369–375.
- Neuer, H., & von Sengbusch, R.: Zucher 15: 49-62 (1943)
- Warmke, H.E.: Carnegie Inst. Washington Yrbk 41: 186-189 (1942); ibid., 43: 153-157 (1944)
- Westgaard, M., 1958. The mechanism of sex determination in dioecious plants. Adv Genet 9: 217–281.
- Zhatov, A.E., Genetika 15 (2): 314-319 (1979)
v/c Dad-
-
 3
3
-
 2
2